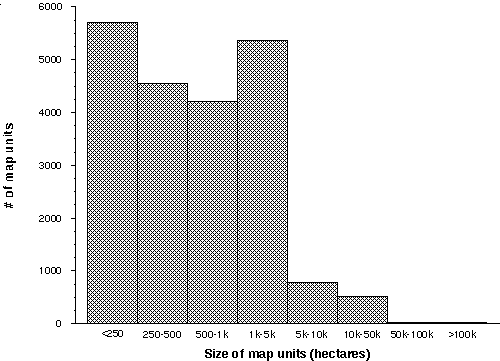What's New
Research & Projects Publications
People
Data
Links
2. LAND-COVER CLASSIFICATION AND MAPPING
Introduction
Methods
Results
Accuracy Assessment
Limitations and Discussion
IntroductionGenerally, the mapping of land-cover is done by adopting or developing a land-cover classification system, delineating areas of relative homogeneity (basic cartographic "objects"), then labeling these areas using categories defined by the classification system. More detailed attributes of the individual areas are added as more information becomes available, and a process of validating both polygon pattern and labels is applied for editing and revising the map. This is done in an iterative fashion, with the results from one step causing re-evaluation of results from another step. For example, the mapping and truthing process may reveal needed corrections to the classification scheme. Finally, an assessment of the overall accuracy of the data is conducted. Where the database is appropriately maintained, the final assessment of accuracy will show where improvements should be made in the next update (Stoms et al. 1994).
In its "coarse filter" approach to conservation biology (e.g., Jenkins 1985, Noss 1987), Gap Analysis relies on maps of dominant natural land cover types as the most fundamental spatial component of the analysis (Scott et al. 1993) for terrestrial environments. For the purposes of GAP, most of the land surface of interest (natural) can be characterized by its dominant vegetation.
Land cover classifications rely on specified attributes, such as the structural features of plants, their floristic composition, or environmental conditions, to consistently differentiate categories (Küchler and Zonneveld 1988). The criteria for a land cover classification system for GAP are: (a) an ability to distinguish areas of different actual dominant vegetation; (b) a utility for modeling vertebrate species habitats; (c) a suitability for use within and among biogeographic regions; (d) an applicability to Landsat Thematic Mapper (TM) imagery for both rendering a base map and from which to extract basic patterns (GAP relies on a wide array of information sources, TM offers a convenient mesoscale base map in addition to being one source of actual land cover information); (e) a framework that can interface with classification systems used by other organizations and nations to the greatest extent possible; and (f) a capability to fit, both categorically and spatially, with classifications of other themes such as agricultural and built environments.
For Gap Analysis, the system that fits best is provisionally referred to as the National Vegetation Classification Standards (NVCS). In recent times, this system has also been referred to as the UNESCO/TNC system (Lins and Kleckner 1996) because it is based on the structural characteristics of vegetation derived by Mueller-Dombois and Ellenberg (1974), adopted by the United Nations Educational, Scientific, and Cultural Organization (UNESCO 1973) and later modified for application to the United States by Driscoll et al. (1983, 1984). The Nature Conservancy and the Natural Heritage Network (Grossman et al. 1994) have been improving upon this system in recent years with partial funding supplied by GAP. The basic assumptions and definitions for this system have been described by Jennings (1993).
At the time the CA-GAP was developing a land-cover map, there was no classification scheme in place that fully satisfied the NVCS. The Nature Conservancy system of Bourgeron and Engelking (1994) for the western United States did not include California. We chose therefore to use the then-current classification used by the California Natural Diversity Data Base (CNDDB) of the Natural Heritage Division in the Department of Fish and Game (Holland 1986) for terrestrial natural communities as a precursor to the NVCS/TNC systems. This classification system of the CNDDB is similar to proposed NVCS alliances in that it is based on dominant species and structure. However, criteria for distinguishing woodland from forest, shrubland from herbaceous, and so on, are less rigorous in the CNDDB system compared to the very precise quantitative criteria in NVCS. Not all CNDDB communities are named for their dominant or indicator species, such as Sierran Mixed Coniferous Forest, but it is unlikely that NVCS will be able to consistently label such complex communities by one or two dominant species either. Despite these limitations and deviations from the NVCS, the CNDDB classification has three strong advantages for use in this generation of GAP. First, the community types are well-described (Holland 1986) and not just listed. Second, the classification scheme is in wide use within federal, state, regional, and local agencies around California and therefore is well-recognized by resource managers. Third, the Natural Heritage Division has assigned state and global ranks to these communities, indicating their rarity and level of threat (Keeler-Wolf 1993). Recently the CNDDB has adopted a new classification system (Sawyer and Keeler-Wolf 1995) which may be more in keeping with the NVCS. Future iterations of CA-GAP may wish to cross-walk the CNDDB classes in our initial land-cover map to this newer system.
Twelve new community types were added to the CNDDB system specifically for CA-GAP, often based on descriptions developed by botanical experts from various parts of the state (Appendix 2-1). Because not all surfaces are naturally vegetated, classes for non-vegetated surfaces such as bare ground, water, and sand dunes, plus human dominated land uses were also defined. The standard for these classes was based on the system developed by the USGS (Anderson et al. 1976) as a hierarchical classification for use in remote sensing applications. The "Anderson" system splits at the first level by land use/land cover type into urban, agriculture, water, and barren land. The second level was disaggregated into several agricultural types (e.g., cropland, orchard, vineyard), water bodies (e.g., rivers, lakes, reservoirs), and various forms of bare ground (salt flats, beaches, sand dunes, bare rock, and quarries or mines). Wetlands were classified according to definitions by Cowardin et al. (1979). By this combination of natural community types and these 27 other land use/land-cover types, all land and water surfaces were classified in the gap analysis database.
MethodsFor CA-GAP, we identified vegetation types by one to three overstory species, each contributing greater than 20% of relative canopy cover. The 20% cover criterion, which we selected to be consistent with the California Vegetation Type Mapping (VTM) survey (Wieslander 1946; see Colwell 1988, for overview), is lower than typically applied to define canopy dominance. For example, the CALVEG classification defines dominant as > 50% (Parker and Matyas 1981). Paysen et al. (1980) define Series based on a single dominant overstory species or genus. The ongoing California Native Plant Society Community Inventory is identifying Series primarily based on a single, overstory dominant, although a few series are based on two co-dominant species, and others are defined by environment (e.g., Alpine Series) (Sawyer and Keeler-Wolf 1995). For our purposes and at our 1:100,000 mapping scale, we found that use of single canopy dominants to type vegetation produced an unacceptable simplification of vegetation composition and pattern. For example, much of the chaparral vegetation in the Southwestern California Region would be mapped as Chamise or Scrub oak chaparral, masking systematic, regional variation in community composition. By using the 20% cover threshold, we retained information on one to three canopy species that are dominant or co-dominant over several-to-many hectares. This area is much larger than plot sizes used in traditional vegetation studies. To avoid confusing these vegetation types with Series or Associations as defined by other systems, we refer to these combinations as Species Assemblages. In the field, species in an assemblage may be uniformly mixed or in a fine mosaic of patches, depending on the scale at which the pattern is observed. This means that in practice, species assemblages in our database can be a series recognized by existing classification systems, a combination of two or three recognized series, or previously unrecognized species combinations.
A map of actual vegetation was produced using summer 1990 Landsat Thematic Mapper (TM) satellite imagery, 1990 high altitude color infrared photography (1:58,000 scale), VTM maps based on field surveys conducted between 1928 and 1940, and miscellaneous recent vegetation maps and ground surveys. Details of the mapping process are provided in Davis et al. (1995), and are only summarized here.
We did not have the resources to map individual stands of homogeneous vegetation. Instead, we have attempted to delimit "landscapes," which we defined as areas of one to many square kilometers in extent with uniform climate, physiography, substrate and disturbance regime, and covered by a single species assemblage or by a mosaic of a few species assemblages associated with different sites (e.g., riparian zones, mesic slopes, xeric slopes). Landscape boundaries were mapped subjectively by photointerpretation of patterns in the satellite imagery. Final delineation of a landscape unit was an iterative process based on evidence from the satellite imagery, 1990 air photos, existing vegetation maps and field reconnaissance. The map was produced using a minimum mapping unit of 100 ha (1 km²). The state was mapped into greater than 21,000 landscape units, or polygons, with a mean size of approximately 1,950 ha.
TM imagery was resampled to the Albers equal-area projection with 100 meter resolution (i.e. 1 hectare pixels), and a false color composite of red, near-infrared and mid-infrared reflectance images was displayed on a video monitor. Obvious landscape boundaries were digitally drafted over the imagery based on image tone and texture. Ancillary information, especially air photos and VTM maps, was used to capture additional compositional changes in vegetation that were not visually obvious in the TM imagery. VTM maps were used to position landscape boundaries on vegetation gradients where no obvious break was visible on either the satellite imagery or in air photos. Two hundred and thirty polygons (excluding urban and agricultural areas) were checked in the field, primarily by roadside reconnaissance.
Using these various sources, a large amount of information was collected for each landscape unit (Table 2-1). Based on our concept of landscape, we recorded a primary species assemblage, which was the most widespread vegetation type or land use/land cover type in the polygon, a secondary and tertiary type, and the fraction of the landscape covered by each type. We also recorded the most widespread wetland assemblage, which was usually riparian vegetation. Each species assemblage was defined by up to three dominant species. We also recorded the occurrence of minor overstory species of special conservation concern (e.g., Juglans californica, Quercus engelmanii, Cupressus forbesii).
Species data were derived from field survey, air photos or from the VTM and other detailed maps. VTM information was used for areas where air photos provided no evidence of recent disturbance, based on the assumption that canopy dominants observed by VTM field crews have not changed over the past 50-60 years. We realize this is a tenuous assumption. We found during our field surveys that the assumption is usually valid for forest and hard chaparral types. Although the relative dominance of species may have changed over the interval, species that were mapped as co-dominants by VTM crews in the 1930s are usually still canopy dominants across the same landscape. The composition of shrubland and grassland types is not as stable over the same interval, and we made special efforts to view these types in the field or to find more current maps. Our landscape units are many square kilometers in extent, and canopy composition can vary greatly from site to site within a landscape. Thus the species assemblages that we have mapped record those species that most frequently dominate most sites in that landscape.
Table 2-1. Landscape attributes for CA-GAP land-cover map units.
Polygon ID Number
Primary Vegetation
Dominant Species 1
Codominant Species 2
Codominant Species 3
Canopy Closure (4 classes)
Fraction of polygon occupied by primary type (10% intervals)
CNDDB Community type
WHR habitat type
Secondary Vegetation
Dominant Species 1
Co-dominant Species 2
Codominant Species 3
Canopy Closure
Fraction of polygon occupied by type
CNDDB Community type
WHR habitat type
Tertiary Vegetation
Dominant Species 1
Co-dominant Species 2
Codominant Species 3
Canopy Closure
Fraction of polygon occupied by type
CNDDB Community type
WHR habitat type
Presence/Absence of 9 wetland habitat types (CA WHR types)
Primary Wetland Vegetation
Dominant Species 1
Codominant Species 2
Codominant Species 3
Presence of canopy species of special status (narrow endemics, RTE species)We have tried to account for fire dynamics by recording recent burns and by retaining information on the pre-burn dominants (e.g., an area of recently burned chamise chaparral that is presently dominated by herbs would be recorded as sparse chamise canopy co-dominated by annual herbs).
Rather than a multi-colored vegetation map, the information we have developed is better treated as a vegetation database linked to a set of areas. One can retrieve distribution data on individual species, unique combinations of species, or vegetation types defined by physiognomy and/or composition (Stoms et al. 1992, Davis et al. 1995). Although the database approach provides a more flexible framework for representing vegetational variation than the traditional vegetation map, it does not eliminate the need for classification in order to simplify and communicate results. We recorded many thousands of unique species (or species/landuse) combinations in over 21,000 polygons. Many unusual species combinations occurred at the margins of regions in transitional environments. We summarize distribution data for individual dominant species and based on plant communities as defined in the California NDDB (Holland 1986), which we derived from the database by an equivalence table assigning each species combination to a unique NDDB community. The criteria for class assignment in the NDDB classification system are qualitative and often not explicitly based on dominant overstory species. Where ambiguities existed, we assigned species combinations to more general types. For example, Holland (1986) identified several pinyon and/or juniper community types in the Mojave Desert region that we aggregated into a single type at his next higher level of classification.
Several procedures were implemented to ensure quality in the final map. The distribution of each dominant plant species in the coverage was compared to the documented distribution recorded in the CalFlora database (see http://s27w007.pswfs.gov/calflora/) which was derived from the Munz flora and revised with some more current data. Apparent outlying locations of species in the GAP database were re-examined to either confirm that the location was documented in the data source or to change the species code if it appeared to be an incorrect interpretation or a data input error. A similar comparison was made for each community type with the written description in Holland (1986).
Results
Of the entire state, 323,492 km² (79%) was classified as vegetation other than agricultural or urban land uses, water, or bare ground. 13% is mapped as various forms of agriculture, with an additional 4.5% in urban or industrial uses. Mean size of the 21,171 landscape units is 1,930 ha, with a median size of 528 ha. The relative proportions of map units by size class is shown in Figure 2-1.
Figure 2-1. Frequency of the size of map units in the CA-GAP land-cover map.
The statewide land-cover map contains distributional information on 221 different land use/cover classes. Of these, 182 were original CNDDB community types described by Holland (1986) or on the California Department of Fish & Game's August, 1987, updated list. In addition, 12 new community types and 27 land use/land cover types were added specifically for CA-GAP. The database contains distributional information on 96 trees, 237 shrubs, 56 herbaceous species or groups of species where they appear as dominants in the canopy (i.e., at least 20% cover in a community type).
Despite the large number of natural communities mapped for the CA-GAP, eight types are particularly widespread (>10,000 km² each), collectively representing nearly 45% of the vegetated land in California (Table 3-2). These types, in descending order of extent, include Mojave Creosote Bush Scrub (34100), Non-native Grassland (42200), Sierran Mixed Coniferous Forest (84230), Coast Range Mixed Coniferous Forest (84110), Sonoran Creosote Bush Scrub (33100), Mojave Mixed Woody Scrub (34210), Blue Oak Woodland (71140), and Foothill Pine-Oak Woodland (71410). On the other hand, 37 types were mapped over less than 25 km² each. Some of these types are indeed rare, and their distributions may be well-known and depicted accurately in the GAP land-cover database (e.g., Monvero Residual Dunes, Valley Sacaton Grassland). Other types, particularly the wetland and riparian communities, may be more widespread than indicated in Table 2-2, but generally occur in small patches below the resolution of the GAP database.
Table 2-2. Total area and percentage of each land-cover type in California.
Land-Cover Code
Land-Cover Type
Total Mapped Distribution (km²)
% of State
Native Plant Communities
21210
Northern Foredunes
3.4
<0.1
21310
Northern Dune Scrub
106.9
<0.1
21320
Central Dune Scrub
13.6
<0.1
22000
Desert Dunes
2,155.1
0.5
23300
Monvero Residual Dunes
3.0
<0.1
31100
Northern Coastal Bluff Scrub
39.1
<0.1
31200
Southern Coastal Bluff Scrub
22.6
<0.1
32100
Northern (Franciscan) Coastal Scrub
469.0
0.1
32200
Central (Lucian) Coastal Scrub
680.5
0.2
32300
Venturan Coastal Sage Scrub
2,102.1
0.5
32500
Diegan Coastal Sage Scrub
1,316.9
0.3
32600
Diablan Sage Scrub
1,107.3
0.3
32700
Riversidian Sage Scrub
744.3
0.2
33100
Sonoran Creosote Bush Scrub
12,625.1
3.1
33200
Sonoran Desert Mixed Scrub
6,323.9
1.5
34100
Mojave Creosote Bush Scrub
43,711.6
10.7
34210
Mojave Mixed Woody Scrub
10,542.3
2.6
34220
Mojave Mixed Steppe
643.7
0.2
34240
Mojave Mixed Woody and Succulent Scrub
574.6
0.1
34300
Blackbush Scrub
1,796.6
0.4
35100
Great Basin Mixed Scrub
6,391.2
1.6
35110
Salvia dorri/ Chamaebatiaria scrub *
30.4
<0.1
35210
Big Sagebrush Scrub
4,341.2
1.1
35211
Low Sagebrush Scrub *
1,623.8
0.4
35212
Silver Sagebrush Scrub *
431.0
0.1
35213
Black Sagebrush Scrub *
18.2
<0.1
35220
Subalpine Sagebrush Scrub
171.4
<0.1
35400
Rabbitbrush Scrub
419.1
0.1
35500
Cercocarpus ledifolius Woodland *
828.2
0.2
36110
Desert Saltbush Scrub
4,723.2
1.2
36120
Desert Sink Scrub
697.4
0.2
36130
Desert Greasewood Scrub
829.6
0.2
36140
Shadscale Scrub
3,492.3
0.9
36150
Desert Holly Scrub *
1,448.0
0.4
36210
Valley Sink Scrub
250.4
0.1
36220
Valley Saltbush Scrub
1,915.5
0.5
36320
Interior Coast Range Saltbush Scrub
27.7
<0.1
37110
Northern Mixed Chaparral
1,652.4
0.4
37120
Southern Mixed Chaparral
219.4
0.1
37200
Chamise Chaparral
5,381.4
1.3
37300
Red Shank Chaparral
965.6
0.2
37400
Semi-Desert Chaparral
2,462.4
0.6
37510
Mixed Montane Chaparral
1,852.0
0.5
37520
Montane Manzanita Chaparral
1,329.0
0.3
37530
Montane Ceanothus Chaparral
1,083.6
0.3
37541
Shin Oak Brush
202.9
<0.1
37542
Huckleberry Oak Chaparral
287.4
0.1
37550
Bush Chinquapin Chaparral
190.4
<0.1
37610
Mixed Serpentine Chaparral
217.2
0.1
37620
Leather Oak Chaparral
98.4
<0.1
37810
Buck Brush Chaparral
4,196.3
1.0
37820
Blue Brush Chaparral
74.3
<0.1
37830
Ceanothus crassifolius Chaparral
2,092.6
0.5
37840
Ceanothus megacarpus Chaparral
583.0
0.1
37900
Scrub Oak Chaparral
2,162.0
0.5
37A00
Interior Live Oak Chaparral
2,105.1
0.5
37B00
Upper Sonoran Manzanita Chaparral
836.3
0.2
37C10
Northern Maritime Chaparral
2.9
<0.1
37C20
Central Maritime Chaparral
218.0
0.1
37D00
Ione Chaparral
0.8
<0.1
37E00
Mesic North Slope Chaparral
628.3
0.2
37G00
Coastal Sage-Chaparral Scrub
345.4
0.1
39000
Upper Sonoran Subshrub Scrub
375.2
0.1
41000
Coastal Prairie
880.3
0.2
42110
Valley Needlegrass Grassland
7.4
<0.1
42120
Valley Sacaton Grassland
9.1
<0.1
42160
Desert Native Grassland *
300.3
0.1
42200
Non-Native Grassland
27,483.4
6.7
42300
Wildflower Field
7.0
<0.1
43000
Great Basin Grassland
117.9
<0.1
44110
Northern Hardpan Vernal Pool
0.1
<0.1
44120
Northern Claypan Vernal Pool
1.6
<0.1
44131
Northern Basalt Flow Vernal Pool
3.9
<0.1
45100
Montane Meadow
360.5
0.1
45200
Subalpine or Alpine Meadow
285.5
0.1
45310
Alkali Meadow
477.0
0.1
45500
Great Basin Wet Meadow *
197.0
<0.1
46000
Alkali Playa
1,436.5
0.4
47000
Pavement Plain
11.3
<0.1
51110
Sphagnum Bog
1.1
<0.1
52110
Northern Coastal Salt Marsh
84.7
<0.1
52120
Southern Coastal Salt Marsh
11.0
<0.1
52200
Coastal Brackish Marsh
310.8
0.1
52310
Cismontane Alkali Marsh
24.5
<0.1
52320
Transmontane Alkali Marsh
33.8
<0.1
52410
Coastal and Valley Freshwater Marsh
501.3
0.1
52420
Transmontane Freshwater Marsh
305.8
0.1
61110
North Coast Black Cottonwood Riparian Forest
11.9
<0.1
61130
Red Alder Riparian Forest
20.3
<0.1
61210
Central Coast Cottonwood-Sycamore Riparian Forest
77.6
<0.1
61220
Central Coast Live Oak Riparian Forest
29.2
<0.1
61230
Central Coast Arroyo Willow Riparian Forest
42.5
<0.1
61310
Southern Coast Live Oak Riparian Forest
26.0
<0.1
61320
Southern Arroyo Willow Riparian Forest
37.7
<0.1
61330
Southern Cottonwood-Willow Riparian Forest
59.9
<0.1
61410
Great Valley Cottonwood Riparian Forest
330.5
0.1
61420
Great Valley Mixed Riparian Forest
73.3
<0.1
61430
Great Valley Valley Oak Riparian Forest
91.6
<0.1
61510
White Alder Riparian Forest
27.5
<0.1
61520
Aspen Riparian Forest
59.4
<0.1
61530
Montane Black Cottonwood Riparian Forest
25.8
<0.1
61610
Modoc-Great Basin Cottonwood-Willow Riparian Forest
80.3
<0.1
61700
Mojave Riparian Forest
58.8
<0.1
61810
Sonoran Cottonwood-Willow Riparian Forest
2.7
<0.1
61820
Mesquite Bosque
273.9
0.1
62100
Sycamore Alluvial Woodland
11.3
<0.1
62200
Desert Dry Wash Woodland
3,003.7
0.7
62400
Southern Sycamore-Alder Riparian Woodland
18.1
<0.1
63100
North Coast Riparian Scrub
99.3
<0.1
63200
Central Coast Riparian Scrub
2.1
<0.1
63310
Mule Fat Scrub
74.7
<0.1
63320
Southern Willow Scrub
1.6
<0.1
63330
Southern Alluvial Fan Scrub *
13.2
<0.1
63410
Great Valley Willow Scrub
18.9
<0.1
63420
Great Valley Mesquite Scrub
22.6
<0.1
63500
Montane Riparian Scrub
119.9
<0.1
63600
Modoc-Great Basin Riparian Scrub
84.6
<0.1
63700
Mojave Desert Wash Scrub
2.0
<0.1
63810
Tamarisk Scrub
105.1
<0.1
63820
Arrowweed Scrub
13.4
<0.1
71110
Oregon Oak Woodland
2,436.9
0.6
71120
Black Oak Woodland
1,641.4
0.4
71130
Valley Oak Woodland
922.0
0.2
71140
Blue Oak Woodland
10,451.8
2.6
71150
Interior Live Oak Woodland
1,731.6
0.4
71160
Coast Live Oak Woodland
1,172.2
0.3
71170
Alvord Oak Woodland
339.7
0.1
71182
Dense Engelmann Oak Woodland
234.8
0.1
71210
California Walnut Woodland
58.5
<0.1
71310
Open Foothill Pine Woodland
1,495.8
0.4
71321
Serpentine Foothill Pine-Chaparral Woodland
836.7
0.2
71322
Non-Serpentine Foothill Pine Woodland
787.8
0.2
71410
Foothill Pine-Oak Woodland
10,180.5
2.5
71420
Mixed North Slope Cismontane Woodland
1,005.3
0.2
71430
Juniper-Oak Cismontane Woodland
301.8
0.1
71600
Oak-Piñon Woodland *
135.8
<0.1
72100
Great Basin Woodlands
9,157.8
2.2
72200
Mojavean Pinyon and Juniper Woodlands
4,035.6
1.0
72300
Peninsular Pinyon and Juniper Woodlands
480.0
0.1
72400
Cismontane Juniper Woodland and Scrub
31.2
<0.1
73000
Joshua Tree Woodland
123.3
<0.1
81100
Mixed Evergreen Forest
4,646.5
1.1
81200
California Bay Forest
13.2
<0.1
81310
Coast Live Oak Forest
2,193.7
0.5
81320
Canyon Live Oak Forest
1,736.2
0.4
81330
Interior Live Oak Forest
2,708.9
0.7
81340
Black Oak Forest
5,614.9
1.4
81400
Tan-Oak Forest
1,824.7
0.4
81B00
Aspen Forest
187.8
<0.1
82100
Sitka Spruce-Grand Fir Forest
339.3
0.1
82310
Alluvial Redwood Forest
742.0
0.2
82320
Upland Redwood Forest
5,407.6
1.3
82410
Coastal Douglas-Fir-Western Hemlock Forest
32.4
<0.1
82420
Upland Douglas-Fir Forest
344.3
0.1
82500
Port Orford Cedar Forest
5.5
<0.1
83110
Beach Pine Forest
14.0
<0.1
83120
Bishop Pine Forest
180.6
<0.1
83130
Monterey Pine Forest
39.2
<0.1
83161
Mendocino Pygmy Cypress Forest
10.5
<0.1
83210
Knobcone Pine Forest
341.6
3.8
83220
Northern Interior Cypress Forest
182.9
<0.1
83330
Southern Interior Cypress Forest
20.5
<0.1
84110
Coast Range Mixed Coniferous Forest
13,539.2
3.3
84120
Santa Lucia Fir Forest
8.4
<0.1
84130
Coast Range Ponderosa Pine Forest
468.9
0.1
84140
Coulter Pine Forest
405.2
0.1
84150
Bigcone Spruce-Canyon Oak Forest
341.6
0.1
84160
Ultramafic White Pine Forest
22.2
<0.1
84171
Northern Ultramafic Jeffrey Pine Forest
496.3
0.1
84180
Ultramafic Mixed Coniferous Forest
262.2
0.1
84210
Westside Ponderosa Pine Forest
9,750.3
2.4
84220
Eastside Ponderosa Pine Forest
6,382.7
1.6
84230
Sierran Mixed Coniferous Forest
15,637.1
3.8
84240
Sierran White Fir Forest
1,055.8
0.3
84250
Big Tree Forest
171.3
<0.1
84260
Modoc White Fir Forest *
942.7
0.2
85100
Jeffrey Pine Forest
3,257.7
0.8
85120
Red Fir-Western White Pine Forest *
1,598.8
0.4
85210
Jeffrey Pine-Fir Forest
4,308.7
1.1
85310
Red Fir Forest
4,475.7
1.1
85320
Southern California White Fir Forest
38.1
<0.1
85410
Siskiyou Enriched Coniferous Forest
245.1
0.1
85420
Salmon-Scott Enriched Coniferous Forest
1,047.1
0.3
86100
Lodgepole Pine Forest
2,572.3
0.6
86210
Whitebark Pine-Mountain Hemlock Forest
429.4
0.1
86220
Whitebark Pine-Lodgepole Pine Forest
588.2
0.1
86300
Foxtail Pine Forest
238.0
0.1
86400
Bristlecone Pine Forest
99.1
<0.1
86500
Southern California Subalpine Forest
52.4
<0.1
86600
Whitebark Pine Forest
233.3
0.1
86700
Limber Pine Forest
21.0
<0.1
91110
Klamath-Cascade Fell-field
59.0
<0.1
91120
Sierra Nevada Fell-field
122.5
<0.1
94000
Alpine Dwarf Scrub
611.0
0.1
Types Other Than Native Plant Communities
11100
Urban or Built-Up Land
18,352.5
4.5
11200-11213
Agricultural types
52,426.3
12.9
11300
Eucalyptus Groves (exotic)
34.8
<0.1
11401-11402
Conifer Plantations
1,335.8
0.3
11510
Streams and Canals
352.8
0.1
11520
Permanently-flooded Lacustrine Habitat
4,059.3
1.0
11521
Intermittently-flooded Lacustrine Habitat
223.1
0.1
11540
Bays and Estuaries
221.1
0.1
11710
Dry Salt Flat
866.1
0.2
11720
Beaches and Coastal Dunes
73.1
<0.1
11730
Sandy Area Other than Beaches
171.3
<0.1
11740
Bare Exposed Rock
5,875.2
1.4
11750
Strip Mines, Quarries and Gravel Pits
247.4
0.1
11760
Transitional Bare Areas
101.3
<0.1
11770
Mixed Barren Land
651.1
0.2
11780
Mud Flats
154.5
<0.1
Statewide Total - All Lands
408,636
100.0
The statewide land-cover map also contains information on the distribution of 59 wildlife habitat types as described by the California Wildlife-Habitat Relationships System (Mayer and Laudenslayer 1988). Of these, 45 types are native upland or wetland habitats, 11 are various agricultural land uses, one represents all urban areas, and 2 are open water habitats in rivers and lakes. The method for assigning habitat types to map units based on the combinations of dominant plant species is described in detail in Chapter 3 as part of the wildlife distribution modeling. Because the focus of GAP is on inventorying natural ecosystems, agricultural habitats in the database are only indicative of the dominant agricultural use of land. Where data were available that distinguised cultivated types at the level of WHR habitats (e.g., deciduous vs. evergreen orchards), this information was used. In many cases, however, the specific types could not be readily determined and were labeled as a more general category such as Orchard and Vineyard (OVN) or even the most general Cropland (CRP). Also the CWHR classification of agricultural habitats was refined midway through the CA-GAP mapping effort. Data recorded prior to this classification revision were not revised. Therefore, agricultural habitats in southern California are particularly likely to be coded as more general types.
Of the 45 native habitats, 8 account for 46% of the total land area in the state (Table 2-3). The most widespread type is Desert Scrub (over 17%). Annual Grassland is second most common (7.1%). Other common habitats are Alkali Desert Scrub, Ponderosa Pine, Sierran Mixed Conifer, Mixed Chaparral, Blue Oak-Foothill Pine Woodland, and Sagebrush, all with between 4.1 and 3.3 percent of the area of California. The least common types, as mapped by CA-GAP, are Saline Emergent Wetland and Palm Oasis, with less than 0.1% each.
Table 2-3. Total area and percentage of each wildlife habitat type in California.
WHR Code
Wildlife Habitat Type (Mayer and Laudenslayer 1988)
Total Mapped Distribution (km²)
% of State
Native Habitat Types
SCN
Subalpine Conifer
3,891
1.0
RFR
Red Fir
5,787
1.4
LPN
Lodgepole Pine
2,097
0.5
SMC
Sierran Mixed Conifer
14,606
3.6
WFR
White Fir
2,935
0.7
KMC
Klamath Mixed Conifer
5,137
1.3
DFR
Douglas-Fir
4,239
1.0
JPN
Jeffrey Pine
7,980
2.0
PPN
Ponderosa Pine
16,385
4.0
EPN
Eastside Pine
4,690
1.1
RDW
Redwood
6,517
1.6
PJN
Pinyon-Juniper
6,359
1.6
JUN
Juniper
8,210
2.0
ASP
Aspen
249
0.1
CPC
Closed-cone Pine-Cypress
1,233
0.3
MHC
Montane Hardwood-Conifer
10,349
2.5
MHW
Montane Hardwood
11,118
2.7
BOW
Blue Oak Woodland
10,579
2.6
VOW
Valley Oak Woodland
3,130
0.8
COW
Coastal Oak Woodland
4,419
1.1
BOP
Blue Oak-Foothill Pine
14,400
3.5
MRI
Montane Riparian
524
0.1
VRI
Valley-Foothill Riparian
800
0.2
DRI
Desert Riparian
363
0.1
POS
Palm Oasis
7
<0.1
JST
Joshua Tree
379
0.1
ADS
Alpine-Dwarf Shrub
755
0.2
LSG
Low Sage
2,778
0.7
BBR
Bitterbrush
218
0.1
SGB
Sagebrush
13,668
3.3
MCP
Montane Chaparral
3,104
0.8
MCH
Mixed Chaparral
14,300
3.5
CRC
Chamise-Redshank Chaparral
10,498
2.6
CSC
Coastal Scrub
7,598
1.9
DSS
Desert Succulent Shrub
4,459
1.1
DSW
Desert Wash
3,017
0.7
DSC
Desert Scrub
70,343
17.2
ASC
Alkali Desert Scrub
16,885
4.1
AGS
Annual Grassland
28,921
7.1
PGS
Perennial Grassland
477
0.1
WTM
Wet Meadow
672
0.2
FEW
Freshwater Emergent Wetland
1,317
0.3
SEW
Saline Emergent Wetland
136
<0.1
EST
Estuarine
216
0.1
BAR
Barren
8,269
2.0
Developed, Cultivated or Water Habitats
URB
Urban
18,378
4.5
Agricultural types
51,620
12.6
RIV
Riverine
354
0.1
Statewide Total - All Lands
408,636
100.0
Accuracy AssessmentGAP land-cover maps are primarily compiled to answer the fundamental question in gap analysis: what is the current distribution and management status of the nation's major natural land cover types and wildlife habitats? Besides giving a measure of overall reliability of the land cover map for Gap Analysis, the assessment will also need to identify which general classes or which regions of the map do not meet the accuracy objectives for the Gap Analysis Program. Thus the assessment identifies where additional effort will be required when the map is updated. We report the results of the accuracy assessment, believing that the map is the best statewide map currently available.
Gap Analysis land-cover maps have already been used in local and regional conservation planning and will likely be used in coordinated national interagency land cover assessments. The maps should be useful for biogeographic analyses and as input to mesoscale models of climate and ecosystem processes. They may well be used to assess the accuracy of smaller-scale land cover maps. These secondary users also need to know the per class accuracy of the maps. Some users would also benefit from having some measure of accuracy by polygon or geographic area, such as where the map is most reliable. We expect that in general, secondary users will need more details about the accuracy assessment to make appropriate use of the map than will primary users who are more familiar with the data.
The purpose of accuracy assessment is to allow a potential user to determine the map's "fitness for use" for their application. It is impossible for the original cartographer to anticipate all future applications of a land cover map, so the assessment should provide enough information for the user to evaluate fitness for their unique purpose. This can be described as the degree to which the data quality characteristics collectively suit an intended application. The information reported include details on the database's spatial, thematic, and temporal characteristics and their accuracy.
Assessment data are valuable for purposes beyond their immediate application to estimating accuracy of a land cover map. The reference data is therefore made available to other agencies and organizations for use in their own land cover characterization and map accuracy assessments (see Data Use and Availability for access information). The data set will also serve as an important training data source for later updates.
Even though we have reached an endpoint in the mapping process where products are made available to others, the Gap Analysis process should be considered dynamic. We envision that maps will be refined and updated on a regular schedule. The assessment data will be used to refine Gap Analysis maps iteratively by identifying where the land cover map is inaccurate and where more effort is required to bring the maps up to accuracy standards. In addition, the field sampling may identify new classes that were not identified at all during the initial mapping process.
Map accuracy can be assessed in many different ways, most commonly by comparing the map to ground observations for a set of sample "points" (Congalton 1991). This approach is not practical for small scale maps such as ours because of the sampling effort required to determine the actual map class at a point on the ground when the minimum mapping unit is 1 km². The size and limited accessibility of some parts of the study area also pose considerable financial and logistical challenges. For these reasons, we have not conducted a formal assessment of the accuracy of the vegetation database. Instead, we have attempted to provide a qualitative measure of map accuracy through roadside reconnaissance and by comparing our map with recent, detailed vegetation maps that have been prepared for parts of the region. Because our approach varied among regions, additional information on map accuracy is deferred to individual regional summaries in the appendix. In general, our studies indicate that the vegetation database has inaccuracies but is generally in high agreement with other, recent vegetation maps (Stine et al. 1996, Stoms 1996, Franklin and Woodcock 1997). However, it is a highly generalized abstraction of vegetation pattern that can serve only for broad regional assessments and inventories. The database will be maintained by the California Department of Fish and Game, and we fully anticipate that the map will undergo periodic revision based on feedback from botanical experts.
Limitations and Discussion
Gap analysis provides a regional overview of the distribution and ownership profile of major terrestrial plant communities and vertebrate species habitats. It is not a substitute for a detailed biological inventory. Our assessment focuses on floristically (and to some extent, structurally) defined plant community types and wildlife habitat types and does not account for variations in stand age or physical stature within a type. For example, we do not distinguish late seral/old growth forest from younger forest of the same general community type.
The extent and spatial scale of the land-cover map make a formal, statistical analysis of map accuracy impractical for both financial and logistical reasons. As a result, we cannot with confidence place error terms on our estimates of area or management status of plant communities.
The method that we used to map land-cover is not suited to the analysis of most wetland types or other communities that are restricted to very localized environments. The mapping method is well suited to analysis of shrubs and trees, but it provides little or no information on the distribution of herbaceous species. Our analyses assume that the vegetation or habitat types attributed to a map unit (polygon) are dispersed uniformly throughout the unit.
Estimates of area made from maps are very sensitive to map scale and mapping methods. For example, vegetation types that typically occur in small patches may be overlooked or their extent underestimated using a vegetation map with relatively coarse spatial resolution. Our vegetation map is less sensitive to spatial resolution than traditional paper maps because we maintain database records of secondary and tertiary vegetation types that are too fine to map using a 100 ha (247 ac) mmu. The point to remember is that our estimates of the acreage and distributions of species and types may differ considerably from areal estimates and distributions of the same types derived from maps prepared at a finer or coarser resolution.
The static nature of the gap analysis data also limits their utility in assessing conservation risk. Our database provides a snapshot of large regions within which land-cover is very dynamic. Thus the database does not reflect how much of a community type may have been lost due to past human land uses nor what proportion is likely to be converted or degraded in the future.
CA-GAP Home | Overview | Report | Download GIS | CD-ROM | Site Index | National GAPTop of Page Send your comments to: stoms@geog.ucsb.edu
UCSB Biogeography Lab Home